기사의 의료 전문가

새로운 간행물
골관절염 실험 모델
최근 리뷰 : 23.04.2024

모든 iLive 콘텐츠는 의학적으로 검토되거나 가능한 한 사실 정확도를 보장하기 위해 사실 확인됩니다.
우리는 엄격한 소싱 지침을 보유하고 있으며 평판이 좋은 미디어 사이트, 학술 연구 기관 및 가능할 경우 언제든지 의학적으로 검토 된 연구만을 연결할 수 있습니다. 괄호 안의 숫자 ([1], [2] 등)는 클릭 할 수있는 링크입니다.
의 콘텐츠가 정확하지 않거나 구식이거나 의심스러운 경우 Ctrl + Enter를 눌러 선택하십시오.
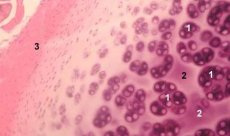
연골은 혈액과 림프관이없는 한 종류의 세포 (연골 세포)만을 함유하는 매우 전문화 된 조직입니다. 연골의 영양은 주로 활액에서의 흡수에 의해 수행됩니다. 연골 세포의 대사는 연골 세포와 주변 조직에 의해 국소 적으로 생성되는 많은 용해 인자에 의해 조절됩니다. 연골 세포의 기능은 또한 세포 외 배지의 조성 (산소 장력, 이온 농도, pH 등), VCM 조성, 세포 및 기질 상호 작용, 물리적 신호에 따라 달라집니다. 실험적 모델링의 주된 임무는 성숙한 세포의 표현형을 바꾸지 않고 세포 외 환경에서 문화를 창조하는 것입니다. 두 번째 과제는 화학 신호 및 / 또는 물리적 신호에 대한 연골 세포의 조기, 지연, 단기 또는 장기 반응을 연구하기위한 문화를 창출하는 것입니다. 연구 체외은 또한 관절염에서 연골 세포의 행동을 연구 할 수있는 기회를 제공한다. 세 번째 과제는 관절에서 다양한 조직의 상호 작용을 연구 할 수있는 공동 치료 시스템 개발입니다. 네 번째 과제는 후속 이식을위한 연골 이식의 준비입니다. 그리고 마지막으로, 다섯 번째 과제는 재생을 자극하고 및 / 또는 연골의 재 흡수를 억제 할 수있는 성장 인자, 사이토 카인 또는 치료제를 연구하는 것이다.
지난 수십 년 동안 단층 세포 배양, 부유 세포 배양, 연골 세포 배양, 이식편 배양, 공 배양, 불멸 세포 배양을 포함하여 관절 연골 세포 배양의 다양한 모델이 만들어졌습니다. 각 문화는 장점과 단점이 있으며 각각은 연골 세포 대사의 한 특정 측면을 연구하는 데 적합합니다. 따라서 연골 조직은 정상 세포 표면 수용체와 정상 세포 - 매트릭스 및 매트릭스 - 세포 상호 작용을 필요로하는 매트릭스 요소의 회전율을 연구하기위한 훌륭한 모델입니다. 동시에, 연골 세포에서의 퇴적물 연구 또는 연골 세포 신진 대사 조절 메커니즘이 격리 된 세포 배양에서 수행되는 것이 좋습니다. 단층의 저밀도 배양은 세포 분화의 과정을 연구하는 데 필요합니다. 천연 또는 합성 매트릭스에 부유 된 배양액은 연골 세포의 기계적 응력에 대한 적응 반응을 분석하기위한 모델이다.
연골 세포 배양
체외 연구를 위해 연골 조직을 선택할 때 몇 가지 중요한 사항을 고려해야합니다. 연골 세포의 매트릭스 구성과 대사 활동은 각기 다른 관절에서 다양하며 후자는 또한 조직 내의 연골 세포의 깊이에 달려 있습니다. 이러한 데이터는 다른 깊이의 연골 영역에서 연골 세포의 고립 된 subpopulations가 연구되었다 여러 실험에서 얻은 것입니다. 관절 연골의 표면과 깊은 층에 위치한 재배 된 연골 세포 사이에 많은 형태 학적 및 생화학 적 차이가 발견되었다. 표면 세포는 드물게 고갈 된 proteoglycan 원 섬유 기질을 합성하는 반면, 더 깊은 세포는 원 섬유와 proteoglycans이 풍부한 기질을 생산합니다. 또한, 표면 세포는 상대적으로 더 작은 비 응집 된 프로 테오 글리 칸 및 히알루 론산을 생성하고,보다 깊게 위치한 연골 세포보다 상대적으로 적은 아그 리칸 및 케 라탄 설페이트를 생성한다. 상이한 깊이의 연골 영역으로부터 분리 된 연골 세포의 신진 대사의 또 다른 중요한 특징은 외인성 자극에 대한 반응이다. M. Aydelotte와 공동 저자에 따르면, 연골의 표층에서 나온 황소 연골 세포는 심부대의 세포보다 IL-1에 더 민감했다.
세포의 행동은 또한 조직의 위치에 달려 있습니다. 이러한 섬유 아세포 성장 인자 (FGF) 및 TGF 베타 같은 성장 인자에 다르게 반응 동일한 동물로부터 취해진 연골 및 귀 가장자리, 연골. FGF는 thymidine, proline과 leucine이 늑골의 연골 세포 배양으로의 결합을 증가 시켰으나, 귀는 증가시키지 않았다. TGF-P는 연골 연골 세포 리브와 귀에 티미 딘의 결합을 증가하지만, 연골 세포 및 프롤린 귀에 티미 딘의 통합에 아무런 영향을 미치지 않았다. 가장 큰 부하를 갖는 구역에서 얻어진 연골 세포는 연골에 대한 부하가 적은 부위의 것과 다르다. 예를 들어, 양 관절 경골 뼈 표면의 중앙부로부터 무릎 관절의 연골 성숙한 연골 세포가 아닌 최대 하중 전달 메 니스 커스에 의해 덮여 생체 내에서 관련성이 밝혀 져야만하지만, 메 니스 커스에 의해 커버되는 영역의 셀보다 더 작은 합성 aggrecan한다. 저자들은 관절의 합성 기능을 조사 할 때 동일한 관절 영역에서 연골을 사용하는 것이 중요하다는 점을 강조합니다.
연골 세포의 신진 대사와 조절 인자에 대한 반응 또한 기증자의 나이, 골격의 발달 및 세포가 취해진 관절의 상태에 따라 크게 좌우됩니다. 인간 연골 세포에서는 증식 반응의 연령에 따라 유의 한 감소가 관찰된다. 가장 큰 감소는 40-50 세 및 60 세 이상의 기증자에게서 관찰됩니다. 또한, 성장 인자 (예 : FGF 및 TGF - 베타)에 대한 증식 반응의 심각성은 노화 동안 감소합니다. 연골 세포의 증식의 양적 변화뿐만 아니라 질적 인 변화도있다. 젊은 기증자 세포 (10-20 세)는 TGF-beta보다 혈소판 유도 성장 인자 (PDGF)에 더 잘 반응하는 반면, 성인 기증자 세포에서는 그 반대가 관찰됩니다. 연골 세포의 합성 기능의 연령 의존적 변화와 성장 인자의 영향에 대한 반응을 설명하기 위해 몇 가지 메커니즘이 사용된다. 그 중에서도 표면 세포 수용체의 수와 친화력 감소, 성장 인자와 사이토 카인의 합성과 생체 활성 변화, 세포 내 수용체 신호의 변형.
관절의 병리학 적 상태는 또한 연골 세포의 형태 및 대사 활동을 변화시킨다. 따라서 J. Kouri와 공동 저자 (1996)는 연골에서 관절염이있는 연골 세포 세 개의 모집단을 확인했다. 연골의 표층과 중층의 연골 세포는 클러스터를 형성하고 더 많은 프로테오글리칸과 콜라겐을 합성합니다. TGF- 베타 및 인슐린 유사 성장 인자 (IGF)는 연골 세포에 의한 프로테오글리칸의 합성을 자극하고 IL-1 및 TNF-α의 효과를 부분적으로 중화시킬 수있다. 연골 절편은 관절염으로 고통되었으며, 관절염 환자의 연골에서 분리 한 연골은 건강한 연골 연골 세포에 비해 TGF 베타의 자극에 더 민감하다. 이러한 차이는 관절 연골의 상층에서 연골 세포의 표현형 변화와 관련이있을 것으로 보인다.
개개의 연골 세포의 분리는 ECM의 단백 분해 효소로 순차적으로 처리함으로써 이루어진다. 분리 된 세포는 ECM으로부터 방출 된 후 de novo 매트릭스 성분의 합성을 연구하는데 이상적 입니다. 일부 저자는 클로스 트리 디움 콜라게나 제만을 사용하고 다른 사람들은 트립신, 프로 나제, DNase 및 / 또는 히알루로니다 제와 연골을 미리 배양합니다. 분리 된 세포의 수는 사용 된 효소에 달려있다. 따라서 처리 할 때, 1g의 콜라겐 조직의 하나 1,4T0 얻을 수 6 4,3-10 - 프로 나제, 및 히알루로게나 사용시 반면, 연골 세포를 6. Collagenase, aggrecan, 단백질, IL-6, IL-8로 처리 할 때 다양한 효소로 순차적으로 처리하는 경우보다 훨씬 더 세포 배양액에 남아있게됩니다. 두 세포 배양액 사이의 차이점에 대해 여러 가지 설명이 있습니다.
- 세포 수용체 손상 또는 효소의 작용에 의해 우울, TGF 베타는 새롭게 절연 연골 세포에서 프로테오글리칸 (1 일)의 DNA 합성을 저해 DNA 및 TGF 베타 자극 단층 배양 된 연골 세포 (7 일)의 프로테오글리칸 합성있다. 그러나 이러한 막 구성 요소를 다시 표현하기 위해서는 실험 시작 전에 적절한 기간이 필요합니다.
- 외인성 proteases는 integrins에 의해 중재 세포와 매트릭스의 상호 작용을 깰 수 있습니다. Integrin 계열은 연골 세포가 VKM 분자에 부착되도록 촉진한다 (Shakibaei M. 외., 1997).이 파열은 기질 유전자의 발현에 영향을 줄 수있다.
- 매트릭스 성분의 잔류 물은 연골 세포의 합성 기능을 조절할 수있다. 인테그린은 ECM의 분해 생성물을 인식 할 수 있기 때문에 단백질 분해 효소에 노출 된 후 조직 복구에 중요한 역할을합니다. T. Larsson과 공동 저자 (1989)는 세포 배양에 손상되지 않았거나 분열 된 프로 테오글리칸을 첨가하면 단백질과 프로테오글리칸의 합성을 촉진한다고보고했다. 그러나, 히알루 론산의 높은 수준은 닭 배아 연골 연골 세포에 의한 설페이트 프로테오글리칸 합성을 포함 상당한 감소 및 돼지 쥐 연골 세포 성숙시킨다. 또한, 히알루 론산 - 심지어는 IL-LB, TNF-A, FGF의 존재하에 세포에서 프로테오글리칸 방출 억제제는, 제는 성장 인자 및 사이토 카인의 생물학적 활성을 중화시키는 것을 나타낸다. 히알루 론산의 작용을 나타내는 정확한 메커니즘은 아직 명확하지 않다. 연골 세포는 세포질의 액틴 필라멘트와 관련된 히알루 론산 수용체를 함유하는 것으로 알려져있다. 히알루 론산과 수용체의 결합은 단백질의 인산화를 자극합니다. 따라서, 이러한 데이터는 막 수용체 세포를 활성화시킴으로써 모체 단백질의 단편화 또는 천연 분자에 의한 연골 세포의 대사 기능의 조절을 입증한다.
- 연골 세포에 의한 기질 단백질의 합성 효소에 의한 신속한 자극은 연골 세포의 형태 변화 및 / 또는 세포 골격의 재구성의 결과 일 수있다.
- 일부 사이토킨 (예 : IL-8)과 성장 인자 (예 : IGF-1, TGF-P)는 ECM에 고정되어 있습니다. 가장 잘 알려진 예는 TGF-β와 decore의 결합으로, 중국 햄스터에서 난소 세포에서 세포 성장을 유도하는 전자의 능력을 감소시킨다. 연령에 따라 연골 장식의 함량이 증가한다는 데이터는 노화에서 TGF-β의 생체 이용률이 감소 함을 나타냅니다. 성장 인자 및 사이토 카인은 배양 중에 기질 잔류 물로부터 방출되어 연골 세포 기능을 조절할 수있다.
연골 세포의 단일 층 배양
분화 된 연골 세포의 표현형은 낮은 수준의 유사 분열 활성뿐만 아니라 II 형 콜라겐 및 조직 특이 적 proteoglycans의 합성에 의해 특징 지어진다. 단층에서 세포가 장기간 재배되고 세포가 여러 번 반복되면 연골 세포가 구형 외형을 잃고 섬유 아세포와 같은 형태를 갖게된다는 증거가 있습니다. 이 섬유 아세포의 증식과 함께 콜라겐 II, IX 및 XI 유형의 합성이 점진적으로 감소하고 콜라겐 I, III 및 Utyopov의 합성이 증가함으로써 세포의 합성 기능이 변형됩니다. 작은 비 응집 proteoglycans은 기능성 aggrecan에 의해 합성됩니다. Synthetzatepsin B와 L은 분화 된 세포에서 매우 낮지 만 분화가 감소하는 과정에서 증가합니다. 콜라게나 제 -1은 분화 된 연골 세포에서 발현되며 장기간 배양되며 발현이 감소하는 반면 metalloproteases (TIMP)의 조직 억제제 생산이 증가합니다.
분화 된 연골 세포는 단층 배양 물에서 부유 세포로 전이 될 때 분화 된 표현형의 콜라겐을 재 발현한다. 분화 과정은 아마 세포의 모양과 관련이 있습니다. 이 특성은자가 연골 세포로 결함 이식을 연구하는 연구자들에 의해 정기적으로 사용됩니다. 생검 물질에서 얻은 소수의 세포를 단층 배양에서 증식시킨 다음 이식하기 전에 3 차원 매트릭스에 다시 놓을 수 있습니다. 아가 로즈 배양으로 옮겨진 탈 분화 된 연골 세포에 의한 특정 표현형의 재 발현은 TGF-β, 오세 인 - 하이드 록시 아파타이트 복합체 및 아스코르브 산으로 자극 될 수있다.
성장 인자 및 사이토 카인의 효과에 반응하여, 연골 세포는 분화 과정 중에 변형된다. 사이토 카인 및 성장 인자에 대한 세포 반응은 미분화 된 및 분화 된 연골 세포간에 상이하다. IL-1은 섬유 아세포의 증식을 자극하는 반면, 미분화 된 연골 세포의 성장은 IL-1에 의해 억제된다. DNA의 합성은 평평하지 않은 연골 세포에서 IGF-1에 의해 자극된다. 분화 된 연골 세포에서 프로 콜라게나 제 생성물에 대한 IL-1β 및 TNF-α의 자극 효과는 미분화 된 연골 세포보다 더 두드러진다.
연골 세포 배양
액체 배지 또는 천연 또는 합성 삼차원 기질에 현탁 된 연골 세포의 배양은 연골 세포의 표현형을 안정화시킨다. 세포는 구형을 유지하고 조직 특이 적 단백질을 합성합니다. 가중 연골 세포 배양은 대개 새로운 세포막 기질 형성의 연구에 권장됩니다. 합성 또는 천연 흡착 중합체의 연골 세포 배양 물은 연골 결함에 세포를 이식하여 관절 연골 조직의 재생을 자극한다. 이식 가능한 세포를위한 합성 또는 자연 환경은 다음과 같은 많은 요구 사항을 충족해야합니다.
- 임플란트는 접착 및 세포 성장을위한 다공성 구조를 가져야하며,
- 중합체 자체 또는 분해 된 생성물은 생체 내 이식 동안 염증 또는 독성 반응을 일으키지 않아야하며 ,
- 이식 담체는 인접한 연골 또는 연골 하골에 결합 할 수 있어야하며,
- 천연 또는 합성 매트릭스는 흡수가 가능해야하며, 그 분해는 조직 재생에 의해 균형을 이루어야하며,
- 연골 회복을 촉진하기 위해, 매트릭스의 화학 구조 및 매트릭스 구조는 연골 세포에 삽입 된 세포 표현형 및 조직 특이 적 단백질의 합성을 유지하는데 도움을 주어야하며,
- 생체 내 이식 하는 동안 , 합성 또는 천연 매트릭스의 기계적 성질을 연구 할 필요가있다.
액상의 연골 세포의 정지
연골 세포의 배양 용액, 메틸 셀룰로스, 아가 로스 겔 (폴리 -2- 하이드 록시 에틸) 또는 콜라겐 오스의 혼합물로 코팅 된 그 벽을 방지 할 수있는 플라스틱 용기에 세포 부착. 이러한 조건 하에서, 연골 세포는 클러스터를 형성하고 주로 aggrecan 및 조직 특이 적 콜라젠 (II, IX, XI 유형)을 합성합니다. 일반적으로 두 가지 유형의 세포가 발견됩니다. 중심에 위치한 세포는 조직 화학 및 초 미세 구조 연구에 의해 확인 된 잘 발달 된 ECM으로 둘러싸인 구형을 유지합니다. 말초 연골 세포는 원반 모양이 둥글며 희귀 한 ECM으로 둘러싸여있다. 그러한 세포의 기능적 특성에 대해서는 알려진 것이 거의 없다.
현탁액으로지지되는 마이크로 담체에 연골 세포를 배양하는 것이 가능하다. 덱스 트란 비즈 (cytodex), 콜라겐 코팅 dextran 비즈 (cytodex III), 콜라겐 타입 I (콜라겐)의 비 공극 microspheres는 microcarriers로 사용됩니다. 이러한 배양 조건 하에서, 연골 세포는 마이크로 캐리어의 표면에 부착되고, 구형을 유지하며, 매트릭스 형 물질을 생성한다. 또한, 콜라겐의 사용은 연골 세포의 증식과 정상적인 표현형의 재발 명을 촉진시킵니다. 따라서, 콜라겐의 미세 구에 대한 연골 세포의 배양은 이식 전에 세포 표현형을 회복 시키는데 사용될 수있다.
액체 배지에서 연골 세포 현탁액을 배양하는 또 다른 방법은 원심 분리에 의해 얻어진 세포 (0.5-1 * 10b ) 로 이루어진 고밀도 비드 형태로 배양하는 것이다 . 이러한 연골 세포는 많은 양의 프로테오글리칸, II 형 콜라겐을 함유하는 매트릭스를 생성 할 수 있지만, 조직 학적, 면역 조직 화학적 및 정량적 방법에 의해 확인되는 I 형 콜라겐은 아니다.
자연 ECM에서 연골 세포의 정지
연골 세포는 3 차원 매트릭스 (부드러운 한천, 아가로 오스, 콜라겐 젤 또는 스폰지, 히알루 론산, 섬유소 접착제, 알지네이트 구슬)로 현탁 배양 될 수 있습니다.
배양 된 아가로 오스 연골 세포는 정상적인 표현형을 유지하고 II 형 콜라겐과 조직 특이 골재 - 새로운 응집체를 합성합니다. 아가로 오스에서 배양 될 때, 세포 합성 proteoglycans은 50 일 동안 배지로 방출된다. 비교를 위해 - 단일 층 배양에서 세포 단계는 배양의 처음 5-6 일 동안 이미 글리코 사 미노 글리 칸으로 과충전된다; 글리코 사 미노 글리 칸의 합성 및 방출 후에 배지에서 배양 될 때 글리코 아미노 글리 칸의 시간 의존적 감소는 처음 8-10 일에 발생한다. 그럼에도 불구하고, 연골 세포가 아가로 오스에서 배양되는 동안의 행동은 생체 내 조건 과 다릅니다 . 아가로 오스에서 많은 수의 합성 Aggregan 응집체가 생체 내 보다 작고 작은 분자를 포함합니다 . TGF-P는 체외 이식편에서 proteoglycans의 합성을 촉진하지만 agarose에서 aggrecan의 합성을 감소시킨다.
알긴산 염은 갈색 해조 유래의 선형 폴리 사카 라이드입니다. Ca 2+ 이온 과 같은 2가 양이온의 존재 하에서 ,이 중합체는 겔이된다. 음으로 하전 된 다당류 매트릭스에 의해 둘러싸여 알긴산에 걸린 각 연골 세포는 기공의 연골의 것과 비슷하다. 두 개의 세그먼트로 이루어진, 알긴산 비드의 연골 형성되는 매트릭스 - 관절 연골 및 기본 조직에서 더 먼 행렬 interterritorial 상당 pericellular 영토 행렬에 대응하는 셀과 연관된 매트릭스 박막. 문화의 30 일에, 상대 및 절대 볼륨이 세포에 의해 점령하고, 알지네이트 비드의 두 부서의 각은 거의 동일 네이티브 연골의 그것입니다. 약 30 일 연골 세포는 구형을 유지하고 관절 연골 콜라겐 II 분자, IX 및 XI 형의 매트릭스 aggrecan 분자 마찬가지이다 aggrecan, 유체 역학적 특성을 생성한다. 동시에, 다른 문화, 현탁 제와 같은 편평 세포 표면에 알긴산 비드는 환경에 직접 방출 및 VCR에 포함되지는 분자 I 형 콜라겐 소량 생성이 존재한다. 알지네이트 비드에서, 연골 세포의 적당한 증식이 관찰된다. 알지네이트 젤 성숙 연골 세포의 배양 8 개월 신진 대사 활동을 잃고 조직 특이 형 콜라겐 II와 aggrecan을 합성하는 것을 계속하지 않는 한.
N. Tanaka와 동료 (1984)는 알긴산 염의 다양한 천연 분자의 확산 특성을 조사한 결과, 70 kD보다 큰 분자는 알지네이트를 통해 확산되지 않는다는 것을 발견했다. 따라서, 알지네이트 중의 세포 배양은 기질 생합성의 조절 및 ECM의 구성을 연구하는데 적합하다. 알지네이트에서 배양 된 세포의 이용 가능성은 전사, 전사 후 및 번역 수준에 대한 펩타이드 조절 인자 및 약리학 적 제제의 효과를 조사 할 수있게한다.
연골 세포는 또한 콜라겐 섬유 I 및 II 유형의 매트릭스에서 배양됩니다. S. Nehrer와 공동 저자 (1997)는 다른 유형의 콜라겐을 함유하는 다공성 콜라겐 - 프로테오글리칸 고분자 매트릭스에서 개 연골 세포의 기능을 비교했다. 그들은 콜라겐 타입 I 및 II를 함유하는 콜라겐 매트릭스에서 배양 된 연골 세포의 생합성 기능의 형태학에서 중요한 차이점을 발견했다. II 형 콜라겐의 매트릭스에있는 세포는 구형 모양으로 감기는 반면, I 형 콜라겐에서는 섬유 아세포와 같은 형태를 가지고있었습니다. 더욱이, II 형 콜라겐의 매트릭스에서, 연골 세포는보다 많은 글리코 사 미노 글리 칸을 생산했다. J. Van Susante et al (1995)은 알지네이트와 콜라겐 (I 형) 겔에서 배양 한 연골 세포의 특성을 비교했다. 저자들은 콜라겐 젤의 세포 수가 현저히 증가한 것을 발견했지만, 배양 6 일부터 세포는 섬유 아세포와 유사한 세포로 변하는 특징적인 표현형을 잃어 버렸다. 알지네이트 겔에서 세포 수의 감소가 관찰되었지만 연골 세포는 정상적인 표현형을 유지했다. 셀당 콜라겐 겔 프로테오글리칸의 양을 알기 네이트에 비해 훨씬 더 높은했지만 알긴산 합성 성장을 계속하는 반면 감소는, 배양 6 일부터 겔 매트릭스 요소 합성에서 관찰되었다.
견고한 3 차원 섬유소 매트릭스는 분화 된 표현형으로 그 안에 부유하는 연골 세포를지지하는 천연 물질입니다. 3 차원 섬유소 매트릭스는 또한 연골 세포 이식을위한 운반체로 사용될 수 있습니다. 섬유소의 장점은 세포 독성의 부재, 공간을 채울 수있는 능력, 접착 능력입니다. 조직 학적 및 생화학 적 연구에 의해, 오토 라디오 그라피 (autoradiography), 전자 현미경, 피브린 겔 내의 연골 세포가 형태를 유지하고, 배양 2 주 후에도 증식하고 매트릭스를 생성하는 것으로 밝혀졌다. 그러나 G. Homminga와 공동 저자 (1993)는 배양 3 일 후에 섬유소의 분해가 시작되고 연골 세포의 탈분화가 진행된다고보고했다.
인공 (합성) ECM에서 연골 세포의 정지
재건 수술 또는 정형 외과 수술을위한 연골 임플란트 는 합성 생체 적합성 매트릭스 에서 체외 에서 분리 된 연골 세포 를 성장시킴으로써 얻을 수 있습니다 .
배양 된 폴리 글리콜 산 연질 세포는 8 주 이내에 정상적인 형태 및 표현형을 증식 및 유지한다. 연골 세포 - 폴리 글리콜 산 복합체는 세포, 글리코 사 미노 글리 칸, 콜라겐으로 구성되어 있으며 외부 콜라겐 캡슐을 가지고 있습니다. 그러나 이러한 임플란트에는 I 및 II의 두 종류의 콜라겐 분자가 있습니다. 연골 세포의 일련의 통로에 의해 탈분화 된 임플란트는 주로 미분화 된 연골 세포로부터의 임플란트에 비해 글리코 사 미노 글리 칸 및 콜라겐의 수가 더 많습니다.
L. Freed와 공동 저자 ( 1,993b) 는 섬유질 폴리 글리콜 릭 산 (HPHC)과 폴리 락트산 (PPLC)에서 인간 및 황소 연골 세포 배양 물의 거동을 비교했다. 저자들은 HSVG 또는 PPLC에서 황색 연골 세포를 6-8 주간 재배 한 후 세포 증식과 연골 기질 재생을 관찰했다. HSBC에서 연골 세포는 구형이었고 연골 기질에 둘러싸인 수족관에 있었다. 8 주간의 체외 배양 후, 재생 된 조직은 건조 물질 (세포 질량의 4 %, 글리코 사 미노 글리 칸의 15 % 및 콜라겐의 31 %)을 50 %까지 함유하고 있었다. PPLK 세포는 스핀들 모양이었고, 소량의 글리코 사 미노 글리 칸과 콜라겐이었다. HSBC에서 세포 성장은 PTCA보다 2 배 더 강했다. 조건 하에서 생체 내에서 1-6 개월 동안 EQAP 및 PPLC 성장 연골 세포 우리는 연골 조직 학적으로 유사한 조직을했다. 임플란트에는 글리코 사 미노 글리 칸, 타입 I 및 타입 II 콜라겐이 포함되어 있습니다.
태아 황소 연골 세포는 다공성의 고밀도 소수성 및 친수성 폴리에틸렌에서 배양되었다. 두 기질 모두에서 7 일간 배양 한 후, 세포는 구형 형태를 유지하였고, 주로 II 형 콜라겐을 함유 하였다. 배양 21 일 후에 친수성 기질이 소수성 기질보다 II 형 콜라겐을 더 많이 포함한다는 것이 밝혀졌다.
연골 조직은 Millicell-CM 필터의 단일 층에서 배양하여 얻을 수도 있습니다. 콘덴서로 필터를 미리 코팅하는 것은 콘드로이친의 부착에 필요합니다. 문화의 조직 검사는 proteoglycans과 타입 II 콜라겐을 함유 한 ECM에서 연골 세포의 축적을 입증합니다. 이러한 배양 물에서 콜라겐 I 형은 검출되지 않습니다. 생성 된 연골 조직의 연골 세포는 구형이지만, 조직 표면에서는 다소 편평 해져있다. 새로 형성된 조직의 두께는 시간이 지남에 따라 증가하였고 세포의 단일 층의 초기 밀도에 의존 하였다. 최적의 배양 조건에서 연골 조직의 두께는 110 μm에 이르렀으며 표면과 깊은 층에서 세포와 콜라겐의 조직은 관절 연골과 유사합니다. VKM에는 콜라겐과 프로테오글리칸이 약 3 배 더 많이 함유되어 있습니다. 2 주간 배양 한 후 matrix-sa의 축적이 관찰되어 필터에서 조직을 추출하여 이식에 사용할 수있었습니다.
Sims 등 (1996)은 폴리에틸렌 옥사이드 - 겔 캡슐화 된 고분자 매트릭스에서 연골 세포의 배양을 연구하여 많은 양의 세포를 주사하여 운반 할 수 있도록 하였다. 무균 쥐의 피하 조직에 주사 한 지 6 주 후에 새로운 연골이 형성되었고 형태 학적으로 유리 연골과 유사한 백색 유분이 특징이었다. 조직 학적 및 생화학 적 연구 결과는 ECM을 생산하는 활발히 증식하는 연골 세포의 존재를 나타냈다.
Explantation
연골 조직의 검사는 아나 및 이화 반응의 과정, 항상성, 흡수 및 수복 과정을 연구하는 데 사용됩니다. 연골 조직 체외 이식편의 연골 세포 는 생체 내 관절 연골과 유사한 ECM의 정상적인 표현형과 구성을지지한다 . 혈청 존재 하에서 5 일간 배양 한 후, 일정 수준의 합성 및 자연 분해가 달성된다. 조직 재 흡수는 여러 가지 제제, 예를 들어 IL-1B, TNF-α, 세균성 리포 폴리 사카 라이드, 레티노 산 유도체 또는 활성 산소 라디칼에 의한 혈청 첨가로 주요 배양 및 배양에서 촉진 될 수 있습니다. 연골의 수복을 연구하기 위해 염증의 용해 매개체 (H 2 O 2, IL-1, TNF-a) 또는 기질의 물리적 파열에 의해 손상이 유도됩니다 .
Organotypic 문화의 방법은 chondrocytes 및 주변 매트릭스에 고립 된 외부 요인의 체외 효과 를 연구하기위한 모델입니다 . 조건 하에서 생체 내에서 연골 세포는 거의 VCR에 위치한되지 않고 서로 접촉하지 않습니다. Explant 관절 연골의 문화는 연골 세포와 주변 세포 외 환경 사이의 특별한 상호 작용뿐만 아니라이 구조적 구성을 유지합니다. 이 모델은 기계적 스트레스, 약리학 적 인자, 성장 인자, 사이토 카인, 호르몬이 연골의 대사에 미치는 영향을 연구하는데도 사용됩니다.
연골 조직 explantation의 또 다른 장점은 proteolytic 효소 또는 세포가 격리 될 때 불가 피한 기계적 요인에 의해 연골 세포 손상의 부재입니다. 수용체 및 기타 막 단백질 및 당 단백질은 손상 요인으로부터 보호됩니다.
연골류의 문화
콘드론은 연골 세포, 그 주변 세포질 및 컴팩트 한 필라멘트 캡슐로 구성된 관절 연골의 구조적, 기능적 및 대사 적 단위이며 매트릭스의 항상성을 담당합니다. 연골은 연골에서 기계적으로 추출되어 몇 차례의 저속 균질화에 의해 수집됩니다. 단일 hondron 트윈 hondrony 다수 (세 이상) 선상 hondrony (열 hondronov) hondronov 정체 : hondrony 연골 네 가지 범주로 나누어 질 수있는 깊이가 다른 구역으로부터 격리.
단일 연골은 보통 손상되지 않은 연골의 중간 층에서 발견되며, 중간과 깊은 층의 경계에서 쌍을 이룬다. 선형으로 위치한 여러 개의 연골은 손상되지 않은 연골의 심층에 전형적이다. 마지막으로, 콘드론의 클러스터는 균질화 후 응집 된 상태를 유지하는 무작위로 조직 된 단일 및 쌍을 이루는 콘드론의 그룹으로 구성됩니다. 연골의 축적은 연골의 큰 단편으로 보통 여러 개의 연골과 방사상으로 위치하는 콜라겐 섬유질을 포함합니다. 즉, 매트릭스의 깊은 층의 전형적인 조직 특징입니다. 연골은 투명 아가로 오스에 고정되어 구조, 분자 구성 및 대사 활동을 연구 할 수 있습니다. Hondron 시스템 - 아가 로스 연골 세포의 전통적인 시스템과 다른 연골의 마이크로 모델로 간주 - 자연 미세 환경을 보존 아가로 오스는 자사의 합성 및 조립을 수행 할 필요가 없습니다. 연골 세포의 문화는 정상 및 병리학 적 조건에서 관절 연골에서 세포와 기질의 상호 작용을 연구하는 모델입니다.
 [22], [23], [24], [25], [26], [27]
[22], [23], [24], [25], [26], [27]
불멸 성 연골 세포의 배양
재조합 DNA 또는 세포 "불멸"할 수 onkogensoderzhaschie 바이러스를 사용하여 영구 세포 라인을 만들 수 있습니다. 불사의 연골 세포는 안정한 표현형을 유지하면서 끝없는 증식 능력을 가지고 있습니다. F. Mallein-게린은 등 (1995)에 종양 유전자 따라서 안정적 콜라겐에게 II, IX 및 XI 유형뿐만 아니라 관절 aggrecan 결합 단백질을 발현 계속 마우스 연골 SV40T 유도 증식 인 것으로 나타났다. 그러나,이 세포주는 단층 배양에서 또는 아가 로스 겔을 배양함으로써, 콜라겐 타입 I 합성하는 능력을 획득한다.
호튼 (W. Horton)과 공동 저자 (1988)는 콜라겐 II 형 mRNA 발현 수준이 낮은 불사의 세포주를 기술했다. 이 세포들은 I-myc 및 y-ra-oncogenes를 함유 한 마우스 레트로 바이러스로 형질 전환시켜 얻어졌다. 이 유형의 세포는 제 2 형 콜라겐의 부재하에 관절 매트릭스의 상호 작용을 연구하고 제 2 형 콜라겐의 합성을 조절하는 독특한 모델입니다.
돌연변이 또는 결실 된 유전자를 가진 연골 동물의 배양은 그들의 생리 기능을 연구하기위한 편리한 모델이다. 이 모델은 연골 매트릭스 조직의 특정 분자의 역할을 공부 연골 대사에 다양한 규제 요인의 영향을 연구에 특히 적합하다. 콜라겐 타입 IX가 피 브릴의 지름을 조절하는 것을 나타내는 정상보다 먼 유전자 합성 형 콜라겐 IX 콜라겐 원 섬유 연골 넓은. I는 제 1 차 일반화 골관절염 가정에서 새로 발견 유전자 돌연변이 COLAI 인코딩 타입 II 콜라겐에 주목한다. 관절 매트릭스 R. Dharmrvaram 등 (1997)이 수행 형질 ( "오염"외래 핵산) 결함 COL 돌연변이 형 콜라겐 II의 영향을 연구하기 위해 2 인간 태아 연골의 AI (시스테인으로 대체 위치 519에 아르기닌) 체외한다.
공동 문화 시스템. 관절에서 연골은 활막, 활액, 인대, 연골 하골에 포함 된 다른 유형의 세포와 상호 작용합니다. 연골 세포의 신진 대사는 이들 세포에 의해 합성 된 다양한 가용성 인자에 의해 영향을받을 수있다. 그래서 관절염 관절 연골은 단백질 분해 효소와 활액 세포에 의해 생성되는 자유 라디칼에 의해 파괴됩니다. 따라서 연골 (coculture)이라고 불리는 연조직과 주변 조직 간의 복잡한 상호 작용을 연구하기위한 모델이 개발되었습니다.
S. 콤브 - Gleise 등 (1995)는 셀이 미다 공막을 분리되는 공 배양 시스템 (COSTAR) (0.4 미크론)의 직접적인 접촉없이 두 세포 형 사이에서 교환 할 수있는 배양 토끼 연골 세포와 조골 세포였다. 이 연구는 수용성 매개체에 의해 연골 세포의 성장을 자극하는 조골 세포의 능력을 보여 주었다.
A.M. Malfait과 공동 저자 (1994)는 말초 혈액 단핵구와 연골 세포 사이의 관계를 조사했다. 이 모델은 염증성 관절염 (류마티스 성 관절염, 혈청 음성 성 척추염 등)에서 사이토 카인이 중재하는 과정을 연구하는데 편리합니다. 모델의 저자는 직경 0.4 μm의 공극을 가진 단백질 결합 막으로 세포를 분리했다. 연구는 리포 다당류 자극 된 단핵 세포는 iFNO IL-1-A, aggrecan 연골의 합성을 억제하여 이미 합성 aggrecan 응집체의 분해에 기여한 것으로 잘되었다.
Kada Tada와 공동 저자 (1994)는 콜라겐 (I 형) 겔의 내피 세포가 연골 세포에서 0.4 μm의 공극 크기의 필터로 외부 챔버와 분리 된 내부 챔버에 배치 된 공동 배양 모델을 만들었습니다. 외부 챔버로부터 완전히 분리 된 상태에서, 인간 내피 세포는 EGF 또는 TGF-α의 존재하에 콜라겐 겔에서 튜브를 형성 하였다. 두 유형의 TGF 세포의 동시 배양으로, 내피 세포에 의한 튜브의 의존적 인 형성이 억제되었다. 이 과정의 연골 세포 억제는 항 -TGF- 베타 항체에 의해 부분적으로 제거되었다. 연골 세포에 의해 생성 된 TGF- 베타가 연골 자체의 혈관 형성을 억제한다고 추정 할 수있다.
S. Groot과 공동 저자 (1994)는 16 일된 태아 생쥐의 뼈의 비대 및 증식 영역에서 연골 세포를 동시에 배양하여 뇌 조직 조각을 만들었다. 배양 4 일 후, 연골 세포의 골아 세포로의 분화 및 골 형성의 개시가 관찰되었다. 배양 11 일 후, 연골의 일부가 뼈 조직으로 대체되었고 골 기질은 부분적으로 석회화되었다. 뇌 조직에 의해 생성 된 일부 신경 펩티드 및 신경 전달 물질은 골아 세포의 신진 대사에 영향을 주거나 수용체를 가지고 있습니다. 그 중에서도 노르 에피네프린, 혈관 활동 장 펩티드, 칼시토닌 유전자와 관련된 펩타이드, 물질 P 및 소마토스타틴을 분리 할 수 있습니다 . 연골 세포와 함께 배양되면 뇌 조직 조각이 연골 세포로의 골아 세포로의 분화 과정을 유도 할 수있는 몇 가지 요인을 생성 할 수 있습니다.
[28], [29], [30], [31], [32], [33]
연골 세포의 배양에 대한 외부 요인의 영향
연골 세포의 신진 대사에 미치는 산소 장력의 영향
대부분의 경우, 연골 세포 배양은 대기 중 산소 장력 조건 하에서 발생합니다. 그럼에도 불구하고 생체 내 연골 세포가 저산소 조건 하에서 존재하고 산소 장력이 다른 병리학 적 조건에 따라 다양 하다는 것은 잘 알려져 있다. 성숙 과정 동안, epiphyses의 혈액 공급에 상당한 변화가 관찰됩니다. 혈관 형성은 성장판의 여러 영역에서 다양하기 때문에 성장 장의 산소 장력도 다양합니다. C. Brighton과 R. Heppenstall (1971)은 토끼의 경골 판에서 비대 위 구역의 산소 장력이 주변 연골보다 적음을 보였다. 일부 대사 매개 변수를 측정 한 결과, 연골 세포가 산소 농도의 국소 변화에 신속하게 반응 할 수 있음이 나타났습니다. 우선, 낮은 산소 장력으로, 연골 세포의 소비가 감소합니다. 산소 장력이 21에서 0.04 %로 감소하면 글루코스 이용률이 증가하고 해당 작용 효소 활성도 및 젖산 합성이 증가합니다. 낮은 산소 장력으로도 ATP, ADP 및 AMP의 절대량은 안정적입니다. 이러한 데이터는 에너지 보존을 최대화하기 위해 연골 세포 대사의 방향성을 나타냅니다. 그럼에도 불구하고, 저산소증의 조건 하에서 합성 활동 및 그에 따른 배상 과정이 바뀐다.
높은 산소 장력은 또한 연골의 매트릭스의 저하, proteoglycans과 DNA의 합성에 감소를 일으키는 연골 세포의 신진 대사에 영향을 미칩니다. 이러한 효과는 원칙적으로 유리 산소 라디칼의 생성을 수반합니다.
연골 세포의 기능에 미치는 이온 농도와 삼투압의 영향
세포 외 매질의 나트륨 함량은 250 - 350 밀리몰, 및 삼투압 - 350-450 mOsm : 천연 연골 이온 농도가 다른 조직과는 크게 다르다. VCR을에서 연골 세포를 분리하고 표준 미디어에서 그들을 잠복기 (DMEM (둘 베코의 최소 필수 중간 - 둘 베코의 최소 필수 매체) 삼투압 - 250-280,7 mOsm)는 크게 셀의 주변 환경 변경합니다. 또한, 표준 미디어 칼슘 및 칼륨의 농도가 기본 조직보다 상당히 낮다하는 음이온 농도 - 상당히 높다.
배지에 자당을 첨가하면 삼투압이 증가하고 세포질 내 H + 및 칼슘 음이온 의 농도가 일시적으로 세포 내로 증가합니다. 이러한 세포 내 변화는 연골 세포의 분화 및 대사 활동에 영향을 줄 수있다. J. Urban et al. (1993) 은 표준 DMEM 배지에서 2 ~ 4 시간 동안 배양 한 분리 된 연골 세포 와 35 8- 설페이트 및 3 H- 프롤린 의 혼입이 원주 조직에서의 10 %에 불과하다는 것을 발견했다. 새로 분리 된 연골 세포 및 연골 조직의 체외 이식편에서 350-400 몰몰의 세포 외 배지의 삼투압에 따라 합성 강도가 최대가되었다. 또한, 상기 삼투압의 표준 DMEM 배지에 단리 된 세포를 위치시킨 후, 연골 세포의 용적이 30-40 % 증가 하였다. 그러나, 12 ~ 16 시간 동안 비 생리 학적 삼투압 하에서 연골 세포를 배양 할 때, 세포는 새로운 조건에 적응하여 세포 외 배지의 삼투압에 비례하여 생합성의 강도를 감소시킨다.
Borgetti P. 등 (1995) 등의 성장, 세포 형태의 매체의 삼투압 효과 및 돼지 연골의 생합성을 조사 하였다. 저자는 삼투압 mOsm 0.28 및 0.38와 미디어에서 배양 된 연골 세포의 유사 생화학 적 및 형태 학적 특성을 보여 주었다. 배양의 처음 4-6 시간 동안의 배지 0.48 mOsm의 삼투압 이후 세포 증식 및 단백질 합성의 감소가 관찰 되었으나하면 결국 제어 값에 도달 이들 파라미터를 복원 일어났다. 0.58 mOsm의 삼투압 세포와 배지에서 연골 세포를 배양 할 때 생리 강도 증식 프로세스를 지원하는 능력을 잃게 6 일 후 연골 세포의 수는 현저히 감소된다. 매체의 삼투압 (osmolarity)이 0.58 몰몰 일 때, 단백질 합성의 깊은 저해가 관찰된다. 발현 된 손실 특성 형질 전환을 연골 세포로 또한, 배양시 삼투압과 매체는 mOsm 0,28-0,38 연골 세포는 높은 삼투압 농도 (mOsm 0,48-0,58) 세포 형태의 현저한 변화에 생리 표현형을 유지 섬유 아세포 유사 세포 및 매트릭스 프로테오글리칸의 조립 전지 용량의 감소이다. 본 연구의 결과는 세포 외 환경의 제한 삼투압의 변동에 연골 세포 응답을 나타냅니다.
다른 이온의 농도 변화는 연골 세포의 생합성 과정에도 영향을 미칠 수 있습니다. 따라서, 5 mmol (표준 DM DM 배지에서의 농도)에서 10 mmol ( 생체 내 에서의 VKM 농도)에서 칼륨 이온의 농도가 증가함에 따라 35 S (황산) 의 포함도 가 반으로 증가 한다. 0.5 mmol 미만의 칼슘 농도는 성숙한 황소 연골 세포에 의한 콜라겐 생성에 기여하는 반면, 1-2 mmol (표준 DM DM 배지에서의 농도에 해당)의 농도는 콜라겐 합성을 현저히 감소시켰다. 고농도의 칼슘 (2-10 mmol)에서 적당한 생합성 증가가 관찰되었다. 다양한 양이온이 연골 세포의 VKM 단백질에의 부착에 관여한다. 따라서 마그네슘 및 망간 이온은 fibronectin과 II 형 콜라겐에 부착을 제공하는 반면 칼슘 이온은 연골 세포가 단백질에 부착하는 데 관여하지 않습니다. 따라서, 기술 된 연구의 결과는 표준 배지에서 배양 한 연골 세포의 생합성 기능에 대한 배지의 칼륨, 나트륨, 칼슘 및 삼투압의 세포 외 이온 변화의 영향을 나타낸다.
연골 세포의 신진 대사에 미치는 기계적 스트레스의 영향
관절의 고정은 ECM에서 대사 과정의 정상적인 과정을위한 기계적 자극의 필요성을 나타내는 연골의 가역 위축을 유발합니다. 대부분의 경우, 사용 된 세포 배양 모델은 정상 대기압 조건 하에서 존재합니다. M. Wright와 공동 저자 (1996)는 기계적 환경이 연골 세포의 신진 대사에 영향을 미치며, 세포의 반응은 압축 하중의 강도와 빈도에 의존한다는 것을 보여 주었다. 관절 연골의 손상 이식편의 부하 실험 관내에서이 정적 하중 단백질 및 프로테오글리칸 합성의 감소를 입증 동하중 동안이 과정을 자극한다. 연골에 대한 기계적 스트레스의 효과를 실현하기위한 정확한 메커니즘은 복잡하고 아마도 세포 변형, 정수압, 삼투압, 전위 및 표면 세포 수용체와 매트릭스 분자와 관련이 있습니다. 이러한 각 매개 변수의 효과를 연구하려면 하나의 매개 변수를 독립적으로 변경할 수있는 시스템을 만들어야합니다. 예를 들어, 체외 이식편 배양은 세포 변형을 연구하는 데는 적합하지 않지만, 연골 세포의 대사 활성에 대한 압력의 전반적인 효과를 연구하는 데 사용될 수 있습니다. 연골의 압축 변형을 셀에 이르게하고, 또한 수압 구배, 전위, 유체 흐름 변화 물리 행렬에서의 수분 함량, 전기 전하의 농도, 삼투압의 레벨 등의 요인의 발생을 수반. 세포 변형은 아가로 오스 또는 콜라겐 겔에 잠긴 격리 된 연골 세포를 사용하여 연구 할 수 있습니다.
연골 세포의 배양에 대한 기계적 자극의 효과를 연구하기 위해 몇 가지 시스템이 개발되었다. 일부 연구자들은 기체 상을 통해 세포 배양에 압력을 가하는 시스템을 사용합니다. 예를 들어 15 분 동안 낮은 주파수 (Hz에서 0.3)에서 13 kPa로의 대기압 이상의 압력을 사용 JP Veldhuijzen 등 (1979)의 캠프 프로테오글리칸 합성 및 DNA 합성을 저하의 증가를 관찰 하였다. R. 스미스 등 (1996) 등이 일정한 압력이 프로세스에 영향을 미치지 아니하는 반면 4 시간 1 Hz에서 연골 불 정수압 (10 MPa로)의 차 배양의 간헐적 노출, aggrecan 콜라겐 타입 II의 합성의 증가를 초래하는 것이 있었다. 유사한 시스템 M. 라이트 동부 등 (1996)를 사용하여 세포 배양 물에서주기적인 압력 연골과 칼슘의 활성화 세포막의 과분극과 관련된 것으로보고 2+ 의존성 칼륨 채널. 따라서, 순환 압력의 영향은 연골 세포 막에서 스트레칭에 의해 활성화되는 이온 채널에 의해 매개된다. 정수압에 대한 연골 세포의 반응은 세포 배양 조건 및 적용 하중의 빈도에 달려 있습니다. 따라서, 환상 정수압 (5 MPa로)는, 0.05, 0.25 및 0.5 Hz의 주파수에서 연골 세포 단층으로 황산의 혼입을 감소 동안 연골 이식편 증가 0.5 Hz에서 포함 황산 상기 주파수.
M. Bushmann 등 (1992)은 아가 로즈 겔의 연골 세포가 배양 된 손상되지 않은 장기와 같은 방법으로 정적 및 동적 기계적 응력에 반응하여 생합성을 변경한다고보고했다. 저자는 기계적 부하가 연골 세포에서 pH의 감소에 이어과 영양 자극을 생성한다는 것을 발견했다.
기계적 스트레칭 효과는 젤에 담긴 세포 배양에 대해 연구 할 수 있습니다. 스트레칭 힘은 컴퓨터로 제어되는 진공을 사용하여 만들 수 있습니다. 시스템이 어느 정도 진공 상태에있을 때 세포 배양이있는 페트리 접시의 바닥이 일정량만큼 확장되고 컵의 바닥 가장자리에서 변형이 가장 적어 중앙에서 최소화됩니다. 연신은 연골 세포의 배양 접시에서 전송 및 배양됩니다. 이 방법에 Vall 홀름-K. 등 (1995) 등은 배양 된 콜라겐 (유형 II) 겔 연골 세포의 mRNA의 발현이 증가 된 것으로 나타났다 2 -integrina한다. 2 P는 R의 인테그린은 타입 II 콜라겐에 결합 할 수있다. 이것은 액틴 결합 단백질과 상호 작용하여 ECM과 세포 골격을 연결하기 때문에 기계 수용체로 간주됩니다.
연골 세포 신진 대사에 대한 pH의 영향
연골 조직의 간질 액의 pH는 다른 조직보다 더 산성이다. A. Maroudas (1980)는 관절 연골의 pH를 6.9로 결정했다. W. Diamant와 공동 저자 (1966)는 병리학 적 조건에서 pH 5.5를 발견했다. 연골 세포가 낮은 PO2에서 살고있는 것으로 알려져 있으며, 이들 세포의 대사 과정에서 해당 당 대사 (총 포도당 대사의 95 %)의 중요한 역할을 나타냅니다. Glycolysis는 다량의 lactic acid의 생산을 동반합니다.
해당 분해 산물에 의한 환경의 산성화 외에도 매트릭스 성분 자체가 매우 중요합니다. 외 프로테오글리칸에 고정 된 음전하의 수많은 이온 성 조성물을 수정 무료 양이온 (예를 들어, H의 고농도가 +, 나트륨 +, K + ) 및 음이온 농도가 낮은 (예를 들면, O2, NPHS가). 또한 기계적 부하의 영향으로 물이 ECM에서 배출되어 고정 음전하의 농도가 증가하고 매트릭스에 더 많은 양이온이 끌어 당겨집니다. 이것은 세포 내 pH에 영향을 미치는 세포 외 배지의 pH 감소를 동반하여 연골 세포의 신진 대사를 변형시킨다. R. WILKIN 및 (1995) A. 홀 세포 외 및 세포 내 환경 행렬 생합성 격리 소 연골 세포의 pH의 영향을 조사 하였다. 그들은 pH의 감소와 함께 매트릭스 합성의 이중 변형을 관찰했다. PH가 약간 감소 (7.4 <의 pH <7.1) na50 % 혼입 증가 35 S0 4 및 3 깊은 산성화 반면, 연골 세포로 H-프롤린 (PH <7.1)와 비교하여 75 %의 합성을 저해 제어. 암모늄 이온으로 동일한 낮은 pH (6.65)를 생성하면 매트릭스 합성이 20 % 감소합니다. 얻어진 결과는 세포 외 기질 합성 배지의 pH의 변형은 세포 내 배지의 pH의 변화에 의해서만 설명 될 수 없음을 나타낸다. 또한, 연골 세포로는 나 산도를 조절하는 능력을 갖는다 +, H + 기, CA + 의존성 C1 _ -NSOZ -CONVEYORS와 H + / ATP 아제를.
[34], [35], [36], [37], [38], [39], [40]
배양액의 조성이 연골 세포의 대사에 미치는 영향
연골 세포를 배양하기위한 배지는 실험 조건과 일치해야한다. 최근에는 송아지 혈청이 배양 조건을 최적화하는 데 사용되었습니다. 그러나 혈청을 사용할 때 여러 가지 중요한 사항을 고려해야합니다.
- 장기 배양에서 조직 주변으로부터의 세포의 외부 성장,
- 다양한 시리즈의 혈청 조성의 다양성,
- 그 안에 미지 성분의 존재,
- 방해의 위험 증가, 다양한 생물학적 요인이 세포의 대사 활동에 미치는 영향을 연구 한 결과
후자의 예는 쥐의 연골 연골 세포에 대한 EGF의 영향에 대한 연구입니다. EGF는 3 H- 티미 딘 의 도입 및 배양 물 내의 DNA 함량의 증가를 자극했다 . 이 효과는 낮은 혈청 농도 (<1 %)에서 더 두드러졌지만, 고농도 (> 7.5 %)에서는 효과가 사라졌습니다.
송아지 혈청이 풍부한 DMEM에서의 합성 및 분해 수준은 생체 내 조건에 비해 유의하게 증가한다는 것이 잘 알려져 있다. in vivo 와 in vitro 대사의 차이는 활액과 세포가 배양되는 환경의 차이에 의해 야기 될 수 있습니다. D. 리 등 (1997)은 배양 된 연골 세포를 아가 로스 20 % 송아지 혈청 및 활액 동종 통상 다수 풍부 DMEM을 함유하는 영양 배지를 이용하여 수송아지이었다. 배지에서 활액의 존재는 활액의 총량의 80 %까지 proteoglycans의 수를 증가시켰다. 이러한 결과는 배양 활액은 유사한 대사율 유도함을 나타내는 생체 내에서 글리코 사 미노 글리 칸 및 세포 분열의 낮은 수준의 합성을 높은 수준.
(1995) G. Verbruggen 등은 합성되었습니다 35 개 혈청없이 S-arrpeKaHa DMEM에서 아가에서 배양 된 연골 세포가 10 % 송아지 혈청으로 보충 된, DMEM에서 관찰 합성의 20 ~ 30 % 수준이었다. 저자는 범위를 결정하는 IGF-1, IGF-2, TGF-P 또는 혈청이없는 배지에서 감소 된 인슐린 생산 aggrecan. 저자들은 결론 지었다 제어 레벨 39-53%에 aggrecan의 100 NG / ㎖ 인슐린, IGF-1, IGF-2의 합성 부분적으로 감소. 이러한 요인들의 조합으로 인해 시너지 또는 누적 현상이 확인되지 않았습니다. 동시에, 100 NG / ㎖ 인슐린의 존재 TGF-P 10ng의 / ㎖가 기준 레벨의 90 % 이상 aggrecan의 합성을 자극. 마지막으로, 혈청 트랜스페린 단독 또는 인슐린과의 조합은 아그 리칸의 합성에는 영향을 미치지 않았다. 종아리 혈청을 소 혈청 알부민으로 대체 하였을 때, 아그 리칸의 응집체 함량은 유의하게 감소 하였다. 인슐린 배양을위한 배지의 농축, IGF 또는 TGF-P는 부분적으로 세포의 aggrecan 응집체를 생산하는 능력을 회복시켰다. 이 경우 IGF-1과 인슐린은 세포 배양에서 항상성을 유지할 수 있습니다. 10-20 NG / ml의 IGF-1이 보충 된 배지에서 배양 40 일 후, 프로테오글리칸 합성을 동일한 수준으로 유지하거나, 심지어 더 높은 매체를 20 % 송아지 혈청을 함유하는과 비교 하였다. 이화 공정은 0.1 % 알부민 용액이 보충 된 배지에서보다 IGF-1이 보충 된 배지에서 느리게 진행하지만, 다소 빠르게 배지에서 20 % 혈청. 수명이 긴 배양에서 20 ng / ml의 IGF-1은 세포의 안정된 상태를 유지합니다.
(1993), D. 리 등은 배지의 조성물의 효과를 비교 하였다 (DMEM, DMEM + 20 % 송아지 혈청 DMEM + 20 NG / ml의 IGF-1)의 DNA 합성에 이식편 연골, 단층 배양의 배양 물 및 아가로 오스 현탁액 . 때 배양 혈청 저자의 존재 아가로 오스의 큰 농도의 연골 세포의 그룹화하는 경향을 관찰했다. 혈청없이 IGF1 배양 세포가, 아가로 오스 원형을 유지, 소규모 그룹으로 수집,하지만 큰 집계를 형성하지 않았다. 단층에서 DNA 합성은 IGF-1이 보충 된 배지에서보다 혈청 함유 배지에서 현저히 높았다 후자에서의 DNA 합성은 농축되지 않은 환경에서보다 훨씬 높았다. IGF-1, DNA 합성의 차이없이 농축되지 매체 및 매체에 아가로 오스 현탁액에 연골 세포를 배양 할 때. 동시에 혈청 배지에 아가로 오스에서 연골 세포를 배양 슬러리, 방사성 뉴클레오티드의 증가 된 결합을 수반 하였다 (3) 다른 환경에 비해 H 티미 딘.
비타민 C는 콜라겐 섬유질의 안정된 나선형 구조 형성에 관여하는 효소의 활성화에 필요합니다. 아스 코르 비 산과 관련하여 결핍 된 연골 세포는 천천히 분비되는 콜라겐의 하이드 록실 화되지 않은 비 - 나선형 전구체를 합성한다. Ascorbic acid (50 μg / ml)의 도입은 II 형과 IX 형 콜라겐의 수산기를 생성시키고 정상 분비물의 분비를 유발합니다. 비타민 C의 첨가는 proteoglycans의 합성 수준에 영향을 미치지 않았다. 결과적으로, 콜라겐의 분비는 proteoglycans의 분비와는 독립적으로 조절됩니다.