기사의 의료 전문가

새로운 간행물
인간 면역 결핍 바이러스 (HIV)
최근 리뷰 : 23.04.2024

모든 iLive 콘텐츠는 의학적으로 검토되거나 가능한 한 사실 정확도를 보장하기 위해 사실 확인됩니다.
우리는 엄격한 소싱 지침을 보유하고 있으며 평판이 좋은 미디어 사이트, 학술 연구 기관 및 가능할 경우 언제든지 의학적으로 검토 된 연구만을 연결할 수 있습니다. 괄호 안의 숫자 ([1], [2] 등)는 클릭 할 수있는 링크입니다.
의 콘텐츠가 정확하지 않거나 구식이거나 의심스러운 경우 Ctrl + Enter를 눌러 선택하십시오.
후천성 면역 결핍증 증후군은 1981 년 미국에서 여러 젊은이들에게 심한 질병이 건강한 사람들에게 병원성이 없거나 약간 병원성이있는 미생물에 의해 발생했을 때 특수 질병으로 격리되었습니다. 환자의 면역 상태를 조사한 결과 일반적으로 림프구 수와 T- 도우미 수가 급격히 감소한 것으로 나타났습니다. 이 상태를 AIDS (영유아 면역 결핍 증후군 - 후천성 면역 결핍 증후군 또는 에이즈)라고합니다. 감염의 방법 (성 접촉, 혈액 및 그 준비를 통한)은 질병 의 전염성을 나타냅니다 .
이 환자 림프절에서 발견 된 이후 AIDS의 원인 물질은, 그것은 Lymphoadenopathy 관련 바이러스를 LAV라는 1983 독립적으로 프랑스 L. 몽 타니)에서 발견되었다; HTLV-III 바이러스 (영어 인간 T- 림프 트로픽 바이러스 III)를 불렀던 American R. Gallo : 이전에 그는 림프 트로픽 바이러스 I 및 II가 발견되었습니다.
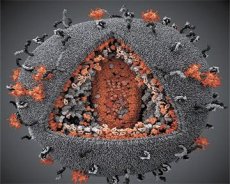
바이러스 LAV와 HTLV-III의 특성의 비교 그렇게 혼동을 피하기 위해, 자신의 정체성을 밝혀, 바이러스가 1986 년이었고, 이름 HIV (영어 인간 면역 결핍 바이러스 -. 인간 면역 결핍 바이러스, 또는 HIV). HIV는 구형이며 직경은 110 nm입니다. 바이러스의 외피는 12 각형과 20 개의 육각형으로 구성된 다면체의 형태를 취합니다. 글리코 실화 된 단백질 gp120의 분자는 각 육각형의 중앙 및 모서리에 위치한다 (숫자 120은 단백질의 분자량을 킬로 돌턴으로 나타낸다). 총 72 개의 gp120 분자가 virion의 표면에 있으며, 각각은 막 내 단백질 gp41과 연관되어 있습니다. 이 단백질들은 이중 지질 층과 함께 비리 온의 수퍼 캡 시드 (막)를 형성합니다.
단백질 gp120 및 gp41은 전구 단백질 Env의 세포 프로테아제 절단 결과로서 형성된다. 단백질 gp41은 척수의 "스터드 (stud)"를 형성하며 세포질 도메인과 막 바로 아래에있는 매트릭스 단백질 p17MA로 연결됩니다. 비리 온의 성숙과 상호 작용하는 분자 p17은 껍질 밑의 20 면체를 형성한다.
Virion의 중앙 부분에서 p24 단백질은 원추형 capsid를 형성합니다. Pb 단백질의 참여로 capsid의 좁은 부분은 virion의 봉투와 연결되어 있습니다. 캡시드 안에는 바이러스 성 게놈 RNA 두 개의 동일한 분자가 있습니다. 그들은 5 '말단에서 뉴 클레오 캡시드 단백질 p7NC와 결합한다. 그들이 형성 비리 온 내로 혼입 게놈 RNA의 분자를 포착으로 그들이 "징크 핑거"라고 -은 시스테인이 풍부한 히스티딘 원자 및 아연을 함유하는은 두 개의 아미노산 잔기 (모티브)이 점에서이 단백질은 흥미 롭다. 캡시드에는 세 가지 효소가 들어 있습니다. Reversase (RT) 또는 pol-complex는 역전사 효소, RNA-ase H, DNA 의존성 DNA 중합 효소를 포함합니다. Revertase는 p66 / p51 heterodimer로 존재합니다. 프로테아제 (PR) - pI는 virion maturation의 과정을 시작하고 실현합니다. 통합 (IN) - p31 또는 엔도 뉴 클레아 제는 숙주 세포의 게놈에 프로 바이러스 DNA가 포함되도록합니다. 캡시드는 또한 씨앗 RNA 분자 (tRNA1 "3)를 함유한다.
세포의 RNA 유전자는 역전사 효소에 의해 9283 염기쌍으로 구성된 DNA 게놈 (DNA-provirus)으로 변환됩니다. 그것은 소위 롱 엔드 반복 또는 LTR (영어 긴 터미널 반복)에 의해 왼쪽과 오른쪽으로 제한됩니다 : 오른쪽에 S'-LTR - 왼쪽과 Z'-LTR -. LTR은 638 염기쌍을 포함하고 있습니다.
HIV 게놈은 9 개의 유전자로 구성되어 있으며, 일부는 중복되어 있고 (몇 개의 독서 틀이 있음) 엑소닌 구조를 가지고 있습니다. 그들은 9 개의 구조 단백질과 6 개의 조절 단백질의 합성을 조절합니다.
바이러스 게놈의 LTR 값은 바이러스 게놈의 기능을 제어하는 다음과 같은 조절 요소를 포함하고 있습니다.
- 전사 신호 (프로모터 영역);
- 폴리 -A의 첨가를위한 신호;
- 신호 캡쳐;
- 신호 통합;
- 양성 조절 신호 (TAT 단백질에 대한 TAR);
- 네거티브 조절 요소 (NEF 단백질에 대한 NRE);
- 3 '말단에서 DNA의 마이너스 사슬을 합성하기위한 시드 RNA (tRNA ™ 3)의 부착 부위; LTR의 5 '말단에있는 신호이며, 이것은 DNA 가닥의 합성을위한 프라이머로 사용됩니다.
또한, LTR은 vRNA 분자를 캡시드 (Psi 요소)로 패킹하는 mRNA 스 플라이 싱의 조절과 관련된 요소를 포함한다. 마지막으로, 긴 mRNA에서 게놈을 전사 할 때, 단백질 합성을 전환시키는 REV 단백질에 대한 두 개의 신호, 즉 조절 단백질에 대한 CAR과 구조 단백질에 대한 CRS가 생성됩니다. REV 단백질이 CAR에 결합하면 구조 단백질이 합성됩니다. 결핍되면 조절 단백질 만 합성된다.
바이러스 게놈의 조절에있어서 다음과 같은 조절 유전자와 그 단백질은 특히 중요한 역할을한다.
- 바이러스의 번식에 대한 양성 조절을 수행하고 조절 TAR 사이트를 통해 작용하는 TAT 단백질;
- 단백질 NEV 및 VPU, NRE 부위를 통한 번식의 음성 대조를 수행함;
- 단백질 REV, 양성 - 음성 대조군을 수행함. Protein REV는 gag, pol, env 유전자의 작용을 조절하고 splicing의 음성 조절을 수행합니다.
따라서 HIV의 번식은 양성, 음성 및 양성 음성의 세 가지 조절하에있게됩니다.
VIF 단백질은 새로 합성 된 바이러스의 감염성을 결정합니다. 그것은 캡시드 단백질 p24에 결합하고 60 분자의 양으로 비리 온에 존재합니다. NEF 단백질은 비리 온에서 소수의 분자 (5-10)에 의해 표현되며 가능하면 외피와 연결됩니다.
VPR 단백질은 세포의 핵에 전송 preintegratsionnyh 단지에 포함 된 G2 단계에서 세포주기를 억제하고 특정 바이러스 성 및 세포 유전자를 활성화 단핵 세포와 대 식세포에서 바이러스 복제의 효율성을 증가시킨다. Virion에서 단백질 VPR, TAT, REV, VPU의 위치는 확립되지 않았다.
자체 단백질 이외에, 비리 온 막의 조성물은 숙주 세포의 일부 단백질을 포함 할 수있다. 단백질 VPU 및 VPR은 바이러스 재생의 조절에 관여합니다.
인간 면역 결핍 바이러스 (HIV)의 항원 변이체
인간 면역 결핍 바이러스 (HIV)는 매우 다양합니다. 한 환자의 유기체에서도 항원 성질이 크게 다른 바이러스의 균주를 분리 할 수 있습니다. 이러한 가변성은 CD4 + 세포의 강력한 파괴와 HIV 감염에 대한 강력한 항체 반응에 의해 촉진됩니다. 서 아프리카의 환자들은 생물학적으로 HIV-1에 가까운 새로운 형태의 HIV를 가지고 있지만 HIV-2와는 면역 학적으로 구분됩니다. 이 바이러스 게놈의 기본 구조의 상 동성은 42 %입니다. DNA-provirus HIV-2는 9671 bp와 그 LTR-854 bp를 포함하고있다. 이후 HIV-2는 다른 지역에서도 분리되었다. HIV-1과 HIV-2 사이에는 교차 면역이 없다. HIV-1의 두 가지 큰 형태가 알려져 있습니다 : O (Outlier)와 M (Major), 후자는 10 개의 아형 (AJ)으로 나뉩니다. 러시아에서는 8 개의 아형이 순환합니다 (AH).
 [
[HIV와 세포의 상호 작용 메커니즘
몸에 침투 한 바이러스는 먼저 특정 CD4 수용체가 들어있는 세포를 공격합니다. 이 수용체는 대 식세포와 단핵구, 특히 바이러스에 민감한 T 조력자에서 많은 수의 T- 조력자를 가지고 있습니다.
인간 면역 결핍 바이러스 (HIV)는 gp120 단백질로 CD4 수용체를 인식합니다. HIV와 세포의 상호 작용 과정은 수용체 매개 흡착 -> 경계 태아 -> 경계 태어난 소포 -> 리소좀에 따라 진행된다. 그것에서, virion 막은 lysosome 막과 합병되고, 핵 캡시드는 supercapsid에서 해방되어 세포질에 들어간다. 핵으로가는 도중에 핵이 파괴되고 게놈 RNA 및 관련 핵심 구성 요소가 방출됩니다. 그런 다음 역전사 효소는 비리 온 RNA에서 DNA의 마이너스 사슬을 합성 한 다음 RNA-ase H가 비리 온 RNA를 파괴하고 바이러스 성 DNA 중합 효소가 DNA의 플러스 가닥을 합성합니다. DNA-provirus의 말단에는 5'-LTR과 3'-LTR이 형성됩니다. DNA-provirus는 잠시 동안 비활성 상태의 핵에 존재할 수 있지만 머지 않아 대상 세포의 염색체에 인테그라 아제의 도움을 받아 통합됩니다. 그것에서, 프로 바이러스는이 T- 림프구가 미생물 항원 또는 다른 면역 대응 세포에 의해 활성화 될 때까지 비활성 상태에있다. 세포 DNA의 전사는 특수 핵 인자 (NF-kB)에 의해 조절된다. 그것은 DNA 결합 단백질이며 T 림프구와 단핵구의 활성화와 증식 동안 대량으로 생산됩니다. 이 단백질은 세포 DNA의 특정 서열과 LTR DNA-provirus의 유사한 서열에 결합하여 세포 DNA와 DNA-provirus의 전사를 유도합니다. DNA-provirus의 전사를 유도함으로써, 그는 바이러스를 비활성 상태에서 활성 상태로, 따라서 지속적인 감염으로 생산적인 상태로 전환시킵니다. 비활성 상태에서 프로 바이러스를 유지하는 것은 매우 오랜 시간 지속될 수 있습니다. 바이러스의 활성화는 세포와의 상호 작용에서 중요한 순간입니다.
바이러스가 세포에 들어간 순간부터 HIV 감염 기간이 시작됩니다 . 바이러스는 10 년 이상 지속될 수 있습니다. 그리고 바이러스의 활성화가 질병 - 에이즈를 시작하기 때문에. 그들의 규제 유전자와 그들의 제품 덕분에, 바이러스는 적극적으로 번식하기 시작합니다. TAT 단백질은 바이러스의 번식 속도를 1000 배로 높일 수 있습니다. 바이러스의 전사는 복잡합니다. 여기에는 전체 길이 및 서브 게놈 mRNA의 형성, mRNA의 스 플라이 싱 및 구조 및 조절 단백질의 추가 합성이 포함됩니다.
구조 단백질의 합성은 다음과 같이 일어납니다. 먼저, 폴리 프로틴 전구체 Pr55Gag가 합성됩니다 (55 kD의 질량을 가진 단백질). - 구조 단백질 P17 각각 형성 매트릭스 (개그 - 폴 다른 단백질 전구체로부터의 samovyrezaetsya) Pr55Gag 바이러스 프로테아제 절단을 초래할있는 (MA) 캡시드 (CA), 뉴 클레오 캡시드 (NC) 및 도메인 PE :이 네 가지 영역을 포함 , p24, p7 및 pb. Polyprotein Pr55Gag의 형성은 바이러스 입자의 형성을위한 주요 조건입니다. 비리 온의 형태 형성 프로그램을 결정하는 것은이 단백질입니다. 그것은 Gag 폴리 프로테인을 원형질막으로 옮기고, 바이러스 입자의 형성과 신진 형성시 단백질 - 단백질 상호 작용과 상호 작용하는 단계를 연속적으로 포함합니다. Pr55Gag는 자유 폴리 리보솜에서 합성됩니다. 단백질 분자는 멤브레인으로 운반되어 그 소수성 패치로 고정됩니다. Gag 단백질의 본래 모양을 창조하기에있는 주요 역할은 CA 영역에 의해한다. NC 도메인 스위치를 형성하는 바이러스 입자로 게놈 RNA의 2 분자 (이하 "징크 핑거"에 의해) 제공한다. 폴리 단백질 분자는 먼저 기질 도메인의 상호 작용으로 인해 이량 체화됩니다. CA와 NC 도메인의 상호 작용의 결과로 이량 체가 6 개 (6 단위)의 복합체로 결합됩니다. 마지막으로, 측면 표면과 연결된 헥사 머 (hexamer)는 미성숙 구형 비 리얼을 형성하며, 그 내부에 NC 도메인에 의해 포획 된 게놈 바이러스 RNA가있다.
다른 전구체 단백질 Prl60Gag - 폴 (m. M. 160 kDa의 가진 단백질) 바로 영역 RB 단백질을 코딩하는 전 영역에 개그 유전자의 번역 Z '말단 중 리보솜의 리딩 프레임을 변화의 결과로 합성된다. 이 Gag-Pol 폴리 프로테인은 Gag 단백질 (1 - 423 아미노산)의 불완전한 서열과 PR, RT 및 IN 도메인을 포함하는 Pol 계열을 포함합니다. Polyprotein Gag-Pol의 분자는 또한 자유 폴리 리보좀에서 합성되어 원 세포막으로 운반됩니다. Polyprotein Prl60Gagpol은 polyprotein Gag와 막 결합 부위에 내재 된 분자간 상호 작용의 모든 부위를 포함합니다. 따라서, Gag-Pol 폴리 프로테인 분자는 멤브레인과 합쳐져 Gag 분자와 함께 개발중인 비리 온에 통합되어 활성 프로테아제를 생성하고 비리 온 숙성 과정이 시작됩니다. HIV-1 프로 테아 제는 이량 체의 형태로만 매우 활성이 있기 때문에 Prl60Gag-Pol의자가 절제를 위해서는이 분자의 이량 체화가 필요합니다. 비리 온의 성숙은 방출 된 활성 프로테아제가 prl60Gag-Pol 및 Gag55를 인식 가능한 부위로 절단 함을 의미한다; 단백질 p17, p24, p7, p6, 리버 타제, 인테그라 아제가 형성되고 바이러스 구조에서 이들의 결합이 일어난다.
봉투 단백질은 글리코 실화, 소포체의 막과 연관된 리보솜에서 합성 된 후, 그것은 gp41 및 gp120의 세포에 프로테아제 절단되고 세포 표면으로 운반된다. 이 경우 gp41은 멤브레인을 투과하여 멤브레인의 내부 표면과 관련된 Gag 단백질 분자의 매트릭스 도메인에 결합합니다. 이 관계는 성숙한 비리 온에서 지속됩니다.
따라서, 바이러스 입자의 어셈블리 전구체 단백질, 숙주 세포의 원형질막에 관련된 RNA 분자, 미성숙 비리 온의 형성 및 세포 표면에서 신진 의해 석방의 응집이다. 출현시, 비리 온은 gp41 및 gp120 분자가 박혀있는 세포막으로 둘러싸여 있습니다. 발아 중 또는 비리 온 성숙 단백질의 방출이 특정 구조 복합체 그들의 연관 성숙 단백질 분해 절단 Pr55Gag 전구체 단백질 Prl60Gag - 폴 바이러스 바이러스 프로테아제되어 이용하여 수행되는, 발생 가능성이 후. 바이러스 형태 형성 과정의 선도적 인 역할은 미성숙 바이러스를 조직하고 조립하는 폴리 프로틴 전구체 Pr55Gag에 의해 수행됩니다. 성숙 과정은 특정 바이러스 성 프로테아제에 의해 완료됩니다.
면역 결핍의 원인
HIV 감염의 면역 결핍의 주요 원인 중 하나는 T- 조력자의 대량 죽음입니다. 다음 이벤트로 인해 발생합니다. 첫째, 바이러스에 감염된 T- 헬퍼 바이러스는 세포 사멸로 사망합니다. AIDS 환자에서 바이러스 복제, 세포 자멸 및 T- 조력자의 수의 감소가 관련되어 있다고 믿어진다. 둘째, T 킬러 세포 (세포의 수십의 일부를 구성 symplasts을 형성 바이러스 나 흡착 gpl20 분자 베어링뿐만 아니라 바이러스에 감염된 바이러스에 감염된 T 헬퍼 세포 (syncytia)에 감염된 T 세포를 인식하고 파괴 바이러스가 번식하여 사망합니다.) 인해 T 헬퍼 세포의 다수의 파괴에 IL-2와 B 림프구 감소 세포막 수용체 발현을 발생 다양한 인터루킨의 방해 합성 (성장 인자와 B 림프구의 분화 -. IL-4, IL-5, IL-6 등) 그 결과 T 킬러 시스템의 기능이 침해 당한다. 보체 및 대 식세포 시스템의 활성 억제가 일어난다. 바이러스에 감염된 대 식세포와 단핵구는 오랫동안 죽지 않지만 몸에서 바이러스를 제거 할 수는 없습니다. 수용체와 구조적 유사성 항원 유기체 어떤 상피 세포를 gpl20 인해 마지막 동작의 넓은 스펙트럼 antiretseptornyh 항체 합성 (trophoblasts는 HIV 이식 전송 중개 수용체 포함). 이러한 항체는 다양한 세포 수용체를 차단할 수 있으며자가 면역 질환으로 질환의 경과를 복잡하게합니다. HIV 감염의 결과는 면역 체계의 모든 주요 부분의 패배입니다. 그러한 환자는 다양한 미생물에 대해 무방비 상태가됩니다. 이것은 기회 감염 및 종 양성 질환의 발달로 이어진다. HIV 감염 환자의 경우 3 가지 유형의 암 위험이 높습니다. 카포시 육종; 암종 (피부암 포함); B- 림프구의 악성 변성으로 인한 B- 세포 림프종. 그러나 HIV에는 림프구뿐만 아니라 신경 영양이 있습니다. 그것은 CNS 세포 (성상 세포)에서 두 수용체 매개 세포 내 이입 및 바이러스에 감염된 lymphoblasts와 성상 세포의 식균 작용에 의해 관통한다. 바이러스가 성상 교세포와 상호 작용할 때 symplast도 형성되어 세포 간 채널을 통한 병원체의 확산을 촉진합니다. 대 식세포와 단핵 세포에서 바이러스가 오랫동안 지속 할 수있다, 그래서 그들은 모든 조직에 침투 할 수있는, 저수지 및 신체의 유통 역할을합니다. 감염된 대 식세포는 중추 신경계에서의 HIV의 이동과 패혈증에 중요한 역할을합니다. 10 %의 환자에서 1 차 임상 증후군은 중추 신경계 손상과 연관되어 치매 (치매)로 나타납니다. 따라서 HIV 감염의 영향을받는 사람들에게는 기회 감염, 종양 질환 및 CNS 손상의 3 가지 질병이 있습니다.
HIV 감염 역학
HIV 감염의 원인은 아픈 사람이나 바이러스 운반자 일뿐입니다. 인간 면역 결핍 바이러스 (HIV)는 혈액, 정액, 자궁 경부에서 발견됩니다. 수유부에서 - 모유로. 감염은 분만 전, 출산 전, 출산 전, 출산 전, 출산 전, 출산 전, 출산 전, 출산 전, 출산 전, 출산 전과 출산 전과 출산 전과 출산 전과 출산 전과 출산 전과 출산 전, 음식, 음료 및 곤충에 의한 바이러스 감염 사례는 알려지지 않았습니다.
약물 중독은 AIDS의 확산에 기여합니다. HIV 감염은 매년 증가하고 있습니다. WHO에 따르면 1980 년부터 2000 년까지 5 천 8 백만 명이 HIV에 감염되었다. 2000 년에만 530 만 명이 전세계에 감염되었고 3 백만 명이 에이즈로 사망했습니다. 러시아에서는 2004 년 1 월 1 일 현재 264,000 명의 HIV 양성 반응자가 등록되어 있습니다. HIV 감염자의 절반은 감염 후 11-12 년 이내에 사망합니다. 2004 년 초, 100,000 명의 러시아 시민 중 약 180 명이 HIV 감염 진단을 받았다. 이 수준에서 2012 년까지 러시아에 HIV에 감염된 사람들의 수는 250-300 만 명에이를 것으로 예상됩니다. HIV 감염과의 싸움의 복잡성은 여러 가지 이유에 달려 있습니다. 첫째, 효과적인 치료 방법과 구체적인 예방책이 없습니다. 둘째, HIV 감염 잠복기는 10 년을 초과 할 수 있습니다. 그 지속 기간은 T- 림프구와 그 염색체에 포함 된 DNA- 프로 바이러스의 활성화 순간에 달려 있습니다. AIDS에 감염된 각 바이러스가 운명에 처하게 될지 또는 질병없이 장기간 바이러스가 유행 할 가능성이 있는지는 아직 명확하지 않습니다. 마지막으로 몇 가지 인간 면역 결핍 바이러스 (HIV-1, HIV-2)가 있으며 그 사이의 항원 성 차이가 교차 면역의 형성을 예방합니다. 원숭이의 면역 결핍 바이러스 (SIV) 검출은 HIV의 기원에 대해 밝혀졌습니다. 게놈을 구성하는 SIO는 HIV와 유사하지만 염기 서열이 크게 다릅니다. HIV-2는 혈청 학적으로 HIV-1과 SIV 사이의 중간 위치를 차지하고 뉴클레오타이드 서열은 SIV에 더 가깝다. 이와 관련하여 VM Zhdanov는 HIV-1, HIV-2 및 SIV 바이러스가 공통 조상에서 유래했다고 제안했습니다. R. Gallo에 따르면, SIV 중 하나가 어떻게 든 인체에 들어갔을 때 HIV-1, HIV-2 및 기타 형태의 돌연변이를 일으킨 수많은 돌연변이를 일으킬 수 있습니다.
HIV 감염의 증상
인간 면역 결핍 바이러스는 질병의 병인이 크게 좌우되는 특정 특징을 특징으로합니다. 이 바이러스는 조절 요소 (활성 단계에서 5 분 이내에 5,000 개의 비리 온이 합성 됨)에 의해 결정되는 매우 높은 번식률을가집니다. 융합 단백질 (gp41)의 존재로 인해 바이러스는 감염된 T-helper와 감염되지 않은 T-helper의 융합으로 인해 광범위한 syncytial 구조를 형성하여 대량 살상을 일으킨다. 대형 분자 gp120 분자는 혈액에서 자유롭게 순환하고 감염되지 않은 T 조력자의 수용체에 결합합니다. 그 결과 T 살해자에 의해 인식되고 파괴됩니다. 이 바이러스는 세포 간 세포 간극을 통해 세포로 전파 될 수 있습니다.이 경우 항체가 쉽게 이용되지 않습니다.
HIV 감염에 대한 임상 적 기준
그들은 적어도 하나 개의 다른 알려진 면역 결핍의 원인의 부재에있는 미성년자의 증상 (암, 선천성 면역 결핍, 심각한 기아, 등등. P.)과 함께 적어도 두 가지 심각한 증상이있는 경우 성인 HIV 수립. 심각한 증상은 다음과 같습니다.
- 체중 감량 10 % 이상;
- 지속 된 발열, 간헐적 또는 지속적인;
- 만성 설사.
경미한 증상 : 지속적인 기침, 일반화 된 피부염, 재발 성 포진, 구강 및 인두 칸디다증, 만성 단순 포진, 일반 림프절 병증. 에이즈 진단은 카포시 육종, 크립토 코카인 수막염, 뉴모시스푸스 폐렴의 존재로 이루어집니다. 질병의 임상 적 그림은 기회 감염에 의해 영향을받습니다.
인간 면역 결핍 바이러스 (HIV)의 배양 방법
HIV-1과 HIV-2는 백혈병 TCV4 림프구에서 얻은 TCB4 림프구 -H9의 한 클론의 세포에서 배양 할 수 있습니다. 동일한 목적으로, 성운 세포의 단일 층 배양 물도 HIV-1이 잘 번식하는 데 사용될 수있다. 동물에서 HIV-1 감염되기 쉬운 침팬지.
외부 환경에서의 바이러스 저항성은 낮습니다. 그는 일광 및 자외선 조사의 영향으로 사망하고 일반적으로 사용되는 소독액으로 처리 한 경우 30 분 동안 80 ℃에서 20-30 분 동안 파괴됩니다. 바이러스 함유 물질을 소독하기 위해서는 가장 높은 저항성을 가진 미생물에 효과적이기 때문에 살균 소독제를 사용해야합니다.
HIV 감염의 실험실 진단
바이러스와 HIV 감염을 진단하는 주요 방법은 효소 면역 측정법입니다. 그러나 gpl20 항체를 gpl20에 대하여 관련 항체 나타나는 신체의 점막 상피 세포를 통해 면역 글로불린의 전송을 수행하는 수용체를 포함하여 인간의 특정 세포 수용체에 대한 구조적 및 항원 유사성을 갖는다는 사실에 기인. 이 경우 IFM의 잘못된 결과가있을 수 있습니다. 따라서, 연구 대상의 모든 양성 반응 혈청을 면역 블 롯법 또는 웨스턴 블 럿팅으로 추가 분석한다. 이 방법은 전기 영동 분리 및 표지 된 항 바이러스 항체로 시험 한 후에 연구해야 할 항체의 확인을 기반으로합니다. 바이러스 학적 방법은 바이러스의 문화가 복잡하기 때문에 거의 사용되지 않습니다. H9 림프구의 클론은 바이러스 성 항원 - 진단 테스트 시스템의 필수 구성 요소를 얻기 위해 사용됩니다. CDR 방법은 바이러스 혈증의 초기 단계에서 이미 바이러스를 검출 할 수있게합니다.
HIV 감염의 치료
역전사 효소 (revertase) 또는 바이러스 성 프로테아제의 활성을 효과적으로 억제하는 약물을 발견하거나 합성하는 것이 필요합니다. 그들은 DNA-provirus의 형성을 방지하고 (또는) 바이러스의 세포 내 증식을 억제합니다. HIV 감염 환자의 치료 현대 전략, 바이러스 단백질 분해 효소 (약물 중 하나)과 revergazu (2 다른 약을) 억제 약물의 결합 사용의 원리에 기초 - 결합 (트리플) 요법. 특히 AZT의 감소 활동, Crixivan Phosphazide 특히 번식의 초기와 후기 단계에 HIV의 복제를 억제 : 러시아에서 HIV에 감염된 사람의 치료를 위해이 개 지역 약물의 병용을 권장합니다.
특정 예방의 문제는 항체의 중요한 생산없이 바이러스 특이 적 세포 독성 림프구에 기초한 효과적인 세포 매개 면역의 형성을 보장하는 백신을 만들 필요가 있다는 것이다. 이러한 면책은 Thl-helpers가 제공합니다. 바이러스 중화를 포함한 항체가 HIV 감염을 억제하는 데 효과적 일뿐만 아니라 높은 수준에서 세포 매개 면역을 억제 할 수도 있습니다. 따라서 항 HIV 백신은 기본적으로 다음 두 가지 기본 요구 사항을 충족해야합니다. A) 절대 안전하고 b) T 세포 독성 림프구의 활성을 자극합니다. 죽은 (불 활성화 된) 바이러스 및 높은 방어 성을 지닌 개별 항원으로부터 얻은 다양한 백신의 효과를 연구합니다. 이러한 항원은 비리 온 자체로부터 분리되거나 화학적으로 합성 될 수있다. 유전 공학적 방법에 기초한 백신이 제안되었다. 강력한 면역 원성을 가진 항원의 합성을 담당하는 HIV 유전자를 가지고있는 재조합 백신 바이러스입니다. 이 백신의 효과에 대한 결정은 HIV 감염의 잠복 기간이 길고 병원균의 높은 가변성 때문에 상당한 시간이 걸립니다. HIV에 대항하여 매우 효과적인 백신을 개발하는 것은 긴급한 근본적인 문제입니다.